布朗克斯沃尔小鼠的较少的GABA能神经元,加剧焦虑和降低高频脑电图组件的遗传性耳聋模型外文翻译资料
2023-01-06 11:27:11
布朗克斯沃尔小鼠的较少的GABA能神经元,加剧焦虑和降低高频脑电图组件的遗传性耳聋模型
关键字:布朗克斯沃尔小鼠,焦虑行为,GABA能中间蛋白,小清蛋白,高频率EEG组件,发育障碍
摘要:突变的bv/bv小鼠的大多数但不是所有的初级听觉感受器,内耳毛细胞和传入神经元退化导致感音性聋。然而,很大程度上不清楚中枢神经系统(CNS)突变的影响。在此研究中,我们进行了行为、 形态和电生理的研究,澄清在bv/bv小鼠的中枢神经系统功能障碍。这些突变小鼠表现出焦虑与正常水平相比明显提高。免疫组化分析显示,bv/bv小鼠前扣带回、听觉皮层中的GABA能神经元显著减少。目前研究结果表明,bv/bv小鼠可能因为大脑的发展中断。此外,bv/bv小鼠相比对照组小鼠,包含高频率范围的gamma;的脑电成分显著减少,这可能干扰皮质的抑制作用。总之,这些结果表明bv/bv小鼠的皮质发展的中间神经元的电生理的组件发生了改变。我们建议建立新的bv/bv小鼠的基因型以便为研究神经元的发育障碍的基本机制提供新的视角。
- 简介
纯合子的布朗克斯沃尔(bv/bv)小鼠患有听力障碍与耳蜗受损(Whitlonet al.,1996;Tucker et al., 1999)。 bv/bv鼠有常染色体的隐性突变,首次由Deol和Gluecksohn-Waelsch (1979年)发现。但据报道,位于小鼠的5号染色体的基因突变是未知的(Bussoli et al.,1997年)。阈值的听性脑干反应(ABR)及畸变产物耳声发射(DPOAE)高程已被报告(Schrott et al., 1989; Schrott et al., 1991; Horner et al., 1985),这表明毛细胞功能障碍的存在。其中一位作者(M.I.)报道,成年bv/bv小鼠的外毛细胞功能波动以ABR和DPOAE进行测试(Inagaki et al., 2006)。另一项研究表明在bv /bv小鼠的下丘神经元的异常反应模式由于除了内毛细胞变性外的毛细胞损伤造成的(Sterbing and Schrott-Fischer,2003)。这些研究结果表明,bv /bv突变影响两种类型的毛细胞在形态和功能水平。
另一只小鼠模型,7和4号染色体上发生隐性突变,结果出现耳聋综合征和盘旋的行为(Rinchik et al., 1991)。因此,盘旋行为据报道是小鼠模型的耳聋行为表现型之一。bv/bv 小鼠标盘旋行为是其特征的行为表型,众所周知,不会在所有的变异小鼠中观察到这种行为(Ishiguro et al.,2007)。石黑浩等人提出的盘旋的行为可能由一种不对称的纹状体功能引起。结果表明,在同侧纹状体中注射多巴胺Drd1受体激动剂A68930(首选的盘旋方向)可以减少旋转(Ishiguro et al.,2007),,这表明在此模型中遗传性耳聋是一种中枢神经系统 (CNS) 异常的圈数引起的。
另一方面,bv /bv小鼠在测试期间处理无盘旋苦苦挣扎的响应的行为(个人通信),这可以解释为高度的焦虑。然而,情感方面,bv /bv小鼠的行为没有受到很好的审查。官崎等人(1985年)报告说,reeler小鼠的大脑、小脑皮质和海马中存在细胞结构的解体,听力赤字基于其异常的 ABR. reeler小鼠的异常ABS记录是由于中央而不是外围的变化(Zheng et al., 2003)。在在范式高架加迷宫测试中,青春期的杂合reeler小鼠表现出显著的较低级别的焦虑相关的行为。同时,缺乏尿皮,促肾上腺皮质激素释放的激素(CRH)的家庭成员的老鼠表现出基础性的耳蜗阈值高以及外毛细胞的听力障碍与功能不良。这些小鼠也证明,十分的焦虑行为由于外侧间隔的CRH 受体 2 (Crhr2)的 mRNA 水平减少(Vetter et al., 2002)。这些报告指出,听力障碍和情感的扰动项可能与小鼠品系有关。在此研究中,我们打算通过比较定量的bv /bv和控制小鼠焦虑行为确认bv/bv小鼠有加剧的焦虑行为。
焦虑状态的行为研究表明,小鼠在alpha;2-gabaa受体(GABR2)的敲点突变后,地西泮未能引起光–黑暗试验和范式高架十字迷宫的抗焦虑活性。这是因为没有观察到小鼠的alpha;3或alpha;5-gabaa受体的突变。这样,焦虑行为的监管可能归因于神经电路的特定的GABAA受体亚型的表达(Louml;w et al., 2000)。GABAA受体的多样性与相应的GABA能中间神经元的多样性(Mouml;hler, 2009)。此外,有趣的是,bv/bv小鼠的新生儿耳蜗的免疫缺乏的GAD在I型径向纤维上(Sobkowicz et al., 1999)。在这项研究中,我们通过计算PV(光伏)含谷氨酸脱羧酶(GAD1;又名GAD67)阳性神经元,进行了免疫组化研究探讨bv/bv突变小鼠的中枢神经系统GABA能中间神经元的形态学方面的内容。
在目前的实验性系统,,如果bv/bv小鼠显示GABA能组织学异常,它将可能是引起神经活动障碍的原因。以前的皮质和海马的研究已经表明,GABA能中间神经元的功能反映在高频(70–500 Hz)脑电图—头颅定位片(EEG)的组成部分称为高gamma;,快速的涟漪(Ylinen et al., 1995; Bragin et al., 2004;Klausberger et al., 2004; Le Van Quyen et al., 2006; Oke et al.,2010) 相关研究表明,高频振荡是由颗粒状的锥体细胞的胞体和树突皮质层上的突触后神经元活动引起的,由GABA能中间神经元形成的驱动的。(Jones et al., 2000)。因此,在当前研究中,我们还比较了bv/bv和对照组小鼠的皮质的高频EEG之间的区别。
2.结果
2.1 行为数据
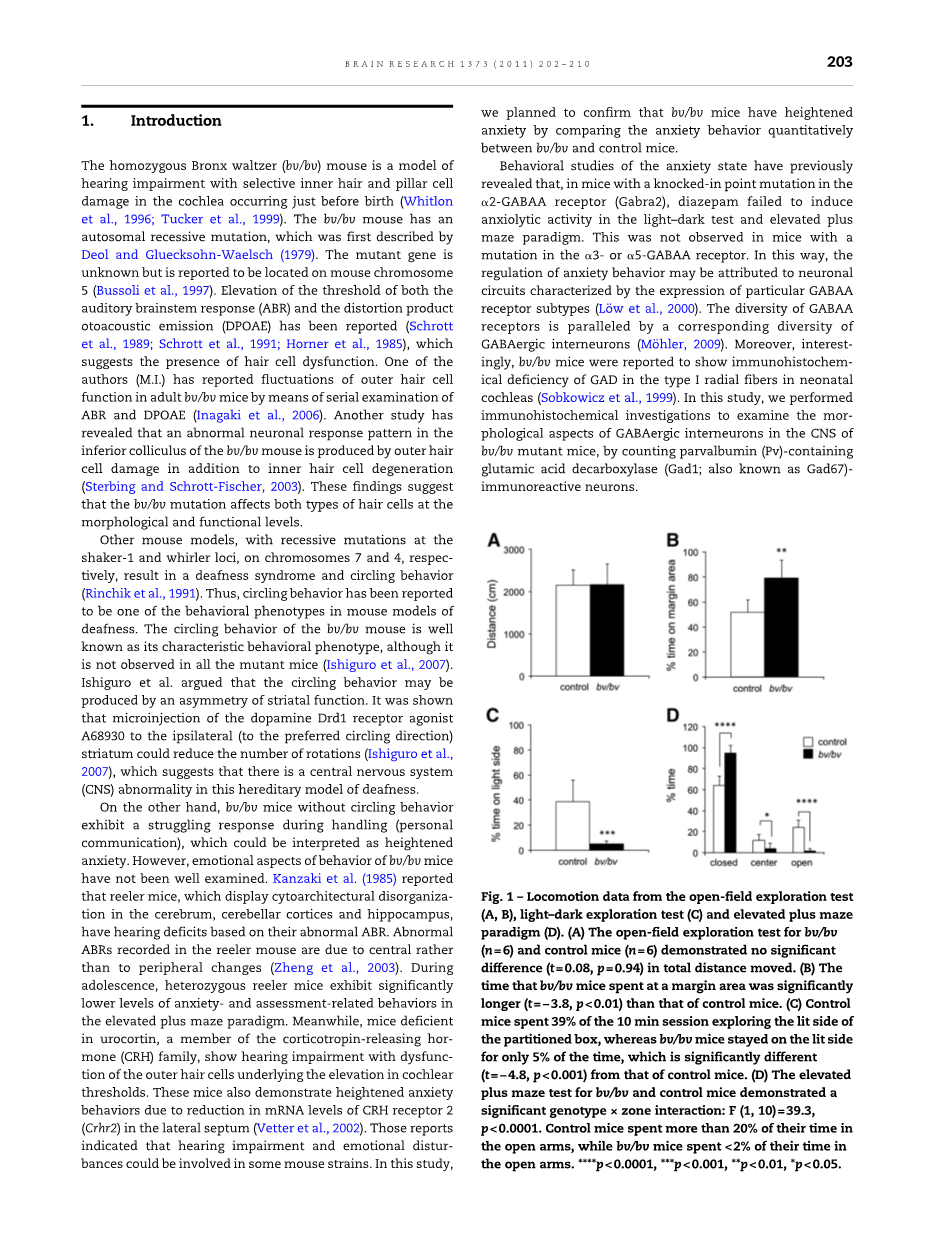
在开放式野外勘探测试,bv/bv和对照组小鼠移动距离相近,分别为2174plusmn;474厘米和2155plusmn;364厘米(图1A)。因此,bv/bv小鼠的广义的自发活动无异常。有趣的是,对照组小鼠在边缘区的时间51.6% ,bv/bv小鼠为80%,其延长时间明显高于对照组[T = -3.8,P lt;0.01]。bv/bv小鼠也没有表现出盘旋的行为。
在明暗勘探测试,bv/bv小鼠在盒子的侧面点燃花费的时间比对照组(39.0%)[T = -4.8,P lt;0.001]的显著缩短5.1% 。
在范式高架十字迷宫测试中,有基因型times;区的相互作用[F =39.3,P lt;0.0001]。
随着事后分析显示,bv/bv小鼠在开放臂[1.8%对24.2%:P lt;0.0001]和中心区域[4.3%对11.7%:P lt;0.05]比没有控制小鼠花费时间明显减少(图1B)。与此相反,bv/bv小鼠在封闭臂花的时间比对照组[93.9%对64.1%:P lt;0.0001]更长,
2.2 bv/bv小鼠皮层组织病理学特征
进行PV和Gad67的双重荧光免疫染色。检测每个前卤水平,在bv/bv小鼠和对照组小鼠的Pv和Gad67免疫阳性细胞在前扣带回、躯体感觉、视觉和听觉的所有层都可以发现。然而,bv/bv小鼠和对照组小鼠[cingulate; Pv: F=80.0, plt;0.0001, Pv/Gad67:F=67.2, plt;0.0001, somatosensory; Pv: F=26.5, plt;0.001, Pv/Gad67: F=26.5, plt;0.001, auditory; Pv: F=10.3, plt;0.05, Pv/Gad67: F=8.6, plt;0.05]相比,在前扣带回(图2A),体感(图2b)和听觉(图2D),PV数和PV / GAD67免疫反应阳性细胞在三前囟水平显著下降( 1.5, 0.5和minus;0.5mm)。另一方面,没有观察到PV数和PV /GAD67免疫反应阳性细胞在视觉皮层显着减少。通过观察发现,这两种基因型的PV,PV/GAD67阳性细胞(图3A)(图3b)在海马CA1区,CA3区和齿状回(DG)(图3c)这三前水平(minus;1.5,minus;minus;2和2.5毫米)的数量相同。同样,基因型在同一水平之间的细胞内的亚核内的杏仁复合体(基底(BLA),杏仁基底内侧核(BMA)和皮质(CO))的PV数和PV/GAD67免疫反应细胞的数量没有表现出显著的差异(图4A–C)。
2.3 分析新皮层的EEG的脑电频率成分
检测过程中bv/bv小鼠和对照组小鼠没有表现出自发的抽搐行为和发作癫痫。EEGs上,所有的bv/bv小鼠的皮层活动的基线显示不同步的异常的双侧棘波放电偶尔出现(图5A)。bv/bv小鼠的尖峰放电是平均每分钟3.25plusmn;2.84. 但是没有对照组在EEGs上显示棘波放电。
从宽频带皮层脑电图活动提取的高频分量(80,500赫兹),显示为图5b基因型谱。在功率谱模式的平均改变表明,bv/bv小鼠与对照组小鼠相比,在80–500赫兹频段有低功率谱。显著区别为高gamma;频(80–120 Hz) [F=10.4, plt;0.01]。相比之下,通过从宽带EEG中观察显示,这两个群体之间的低频分量的频谱功率模式的平均改变(0–30Hz)[p=0.641]没有显着差异,
3.讨论
在目前的研究中,我们进行了行为,免疫组化和电生理测试实验探究bv/bv小鼠的中枢神经系统功能障碍。在行为测试中,在标准的开放领域使用自动语音追踪系统定量的评估控制组的小鼠和bv/bv小鼠的自发活动,这样可以得到客观的行为数据。我们发现bv/bv突变小鼠不能影响一般的运动活动。另一方面,bv/bv小鼠在光暗探测测试和高架十字迷宫测试中表现出高度的焦虑行为,这已被作为测试啮齿动物的焦虑行为一个生理行为工具(Kulkarni and Sharma,1991)以及被用于研究抗焦虑和焦虑的药物(Lister,1987)。这说明bv/bv小鼠在正常的活动水平伴有高度的焦虑(例如无条件的害怕)是他们的行为现象特征。
免疫组化调查显示bv/bv小鼠的大脑皮层的GAD67阳性细胞在海马,杏仁复合体或纹状体未见明显变化时会有光伏发电现象(数据显示)。,虽然很小的数量的新皮层锥体细胞中也含有钙结合蛋白PV (Stichel et al., 1987; Jinno and Kosaka, 2004),它主要在大量的GABA能中间神经元上表达,明显在皮层的枝状,笼状体(大型或者小型)以及网状的笼状体细胞上(Druga, 2009)
大多数含有PV的细胞具有对Gad以及GABA合成酶的强烈的免疫能力。因此,PV可以作为一个整个大脑的相当大的GABA能中间神经元亚群的主标记(Celio, 1986; Schwartz-Bloom and Sah, 2001)。从这些结论中得出,这些bv/bv小鼠的皮质的Pv/Gad67共表达细胞减少的数量可能意味着突变小鼠的GABA能神经元。因此,我们假定,bv/bv突变因为GABA能中间神经元的减少造成皮层神经网络的限制性去抑制。这些发现也许可以明确bv/bv突变造成GABA能中间神经元在发育中迁移。
此外,我们发现 bv/bv小鼠和正常小鼠相比,大脑皮质高频脑电信号的活动明显地减少了。在目前的研究发现,当皮质主细胞被GABAA受体介导抑制性电位的强烈的抑制时(Whittington et al., 1995;Cunningham et al., 2004),大脑皮层gamma;振荡都是由抑制性中间神经元同步放电的引起的(Fries et al.,2007),以前的细胞内记录研究表明大鼠的诱发胡须在体感皮层的快速脉冲装置中爆发火焰和同时记录的快速震荡活动有关,这反映了抑制性突触后电位(Jones et al., 2000)。这些报道表明,通过神经网络的GABA能中间神经元驱动与高频电活动产生关联的可能性。因此,我们推测,在BV / BV小鼠的高频脑电成分减少反映皮层的失调平衡兴奋/抑制活动。
在bv/bv小鼠的皮层神经元的抑制作用降低可能与他们的高度焦虑有关。大多数的抑制性中间神经元表达GABAA受体,主要的快速抑制性神经递质系统在中枢神经系统。鉴于皮质GABA能在bv/bv小鼠皮层的中间神经元的数量减少,这个合理的假设打乱了GABAA受体系统。( Louml;w et al. (2000))。有报道,小鼠的alpha;2 GABAA受体亚单位基因的点突变,在小鼠的焦虑行为测试过程中,皮层和边缘系统的优势是使小鼠保持安定不焦虑,和使小鼠的抗焦虑水平提高。此外,缺乏WFS1基因的小鼠,以及缺乏同源基因导致人类患有Wolfram综合征,表现出alpha;1和alpha;2亚基在皮层GABAA受体显著下调并且增加环境压力。(Raud et al., 2009)。
最近的研究显示,bv/bv小鼠和缺乏尿激酶型纤溶酶原激活物受体的的小鼠有相似的行为/免疫组织化学现象。(Plaur,也称受体)。uPAR minus;/minus; 小鼠在正常活动水平也表现为焦虑增加。形态上,这两种小鼠表现出大脑皮层的额叶-顶叶的特定区域的扣带含光伏GABA能中间神经元减少(Powell et al., 2003; Levitt, 2005; Eagleson et al., 2010). Bae et al. (2010)。这表明这些区域的缺陷因为部分uPAR minus;/minus;小鼠是出生后肝细胞生长因子/散点因子(HGF/SF)生物活性降低。HGF已被确定为一个引导神经元的运动发展的重要的分子线索。(Powell et al., 2001)。在未来,我们计划研究是否bv/bv小鼠的HGF活性降低。
然而,bv/bv小鼠减少的GABA能中间神经元不仅只是可能是因为发展迁移缺陷,也可能因为分化及存活效应。为了在未来解决这个重要问题,我们将需要进行发展研究,其中会在不同发育阶段比较不同基因型的PV/ GAD67阳性细胞。
有趣的是,在一个基因关联研究最近发现,人类基因编码的受体与自闭症风险存在很大关联。(Campbell et al., 20
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 9 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[286324],资料为PDF文档或Word文档,PDF文档可免费转换为Word



