棉花纤维:一个强大的细胞壁纤维素研究单细胞模型外文翻译资料
2023-01-10 16:03:31
棉花纤维:一个强大的细胞壁纤维素研究单细胞模型
摘要:棉纤维是由种子表皮的单细胞扩展而来。当它们经历阶段性分化的时候可以以纯粹的形式被分离,包括伸长时的初生细胞壁和次生壁增厚时的几乎纯的纤维素合成在。这种特性的融合支持在高通量现代实验技术的背景下关于细胞壁和纤维素合成数据的的清晰的解释。本文将总结关于之前从棉花纤维到构建细胞壁基础知识的贡献,并且通过棉花纤维分化描述细胞壁聚合物的活跃变化。。最近使用稳定棉花转换改变棉纤维细胞壁属性以及棉纤维质量的成功案例也将进行讨论。通过病毒诱导基因沉默改变棉花纤维细胞壁的更快速实验方法的未来展望将被评估。
关键词:纤维素 棉花纤维伸长 棉 果胶 过渡阶段 病毒诱导基因沉默 木葡聚糖
正文:
1.棉纤维细胞壁的重要性
棉纤维、高伸长和加厚的单细胞种子的表皮,是世界上最重要的天然纺织纤维。
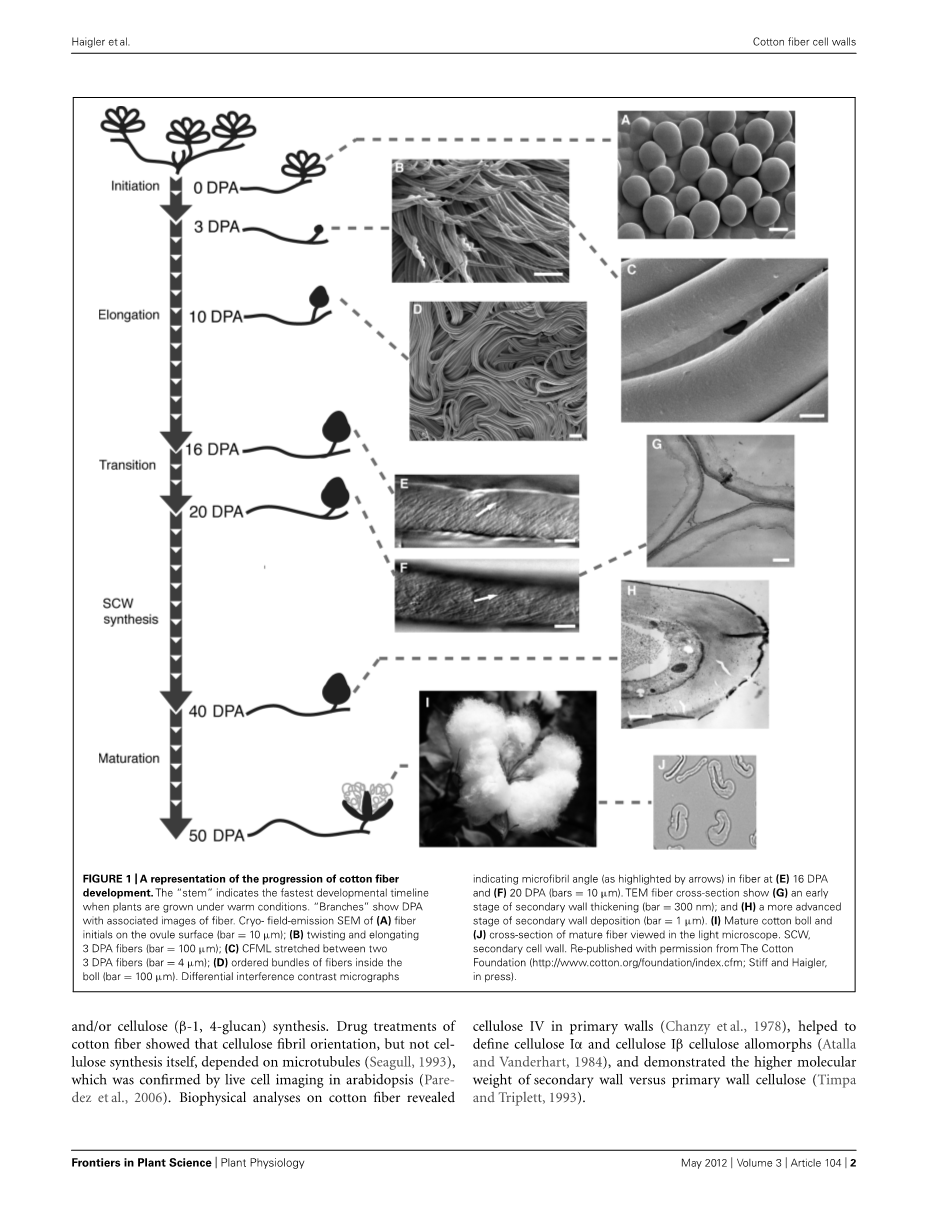
本文将对异源四倍体陆地棉,大多数商业棉纤维的来源,和纤维细胞壁的聚合物,有助于确定棉纤维品质。棉纤维的肿胀开始近花开放的一天的胚珠表面上方(或花)。天开花后(DPA)描述了光纤发展的时间过程,虽然基因型与环境相互作用的影响进展速度(见下文)。纤维正好扩张,这可能涉及细胞壁松弛,由于在选定的表皮细胞膨压增加。在未来的50天棉纤维经历:
- 极限伸长率(超过2.5厘米)通过初生壁的合成;
- 过渡壁增厚和初生壁重塑;
(c)次生壁增厚通过几乎是纯纤维素沉积。
最后,我定义的“成熟”和细胞死亡过程中发生的棉铃打开揭示棉花果在蓬松的纤维(或铃;数据图1)。心皮通常四(或室)一个水果含有约32的种子和~ 500000棉纤维长(皮棉纤维;bowman et al.,2001)。人口的厚壁但短纤维(绒毛纤维和短绒)也存在;这些都是许多工业应用化学纤维素的重要来源。
除了定义形态,棉纤维细胞壁的定义重要的工业纤维质量参数(Wake-lyn et al., 2007). 纤维长度由初生壁的合成是纺纱的本质。纤维强度是由一层薄薄的“线圈”细胞壁层(见下文)和次生壁纤维素,可使纤维的染料深深的吸收水。次生壁纤维素与纤维次生壁增厚周长确定理想的中间程度的量,它的光纤陷入菜豆形状,有利于棉纱线的纺纱要求。纤维素的生物物理性质如结晶度、纤丝角,和聚合度对纤维性能的影响。成熟的棉纤维中含有大于90%(w/w)结晶纤维素,包括纤维次生壁包围的200 nm角化初生壁。
2.棉纤维细胞模型
棉纤维是一种功能强大的细胞壁的研究模型,因为它是一个容易分离的单细胞与细胞壁合成的不同阶段。其他优点包括培养棉花胚珠/纤维的能力(Kim and Triplett, 2001).许多基因在棉花纤维中的表达(Hovav et al.,2008)支持多种细胞壁相关的发育事件。6 DPA后,冰冻的棉纤维可以通过温和的研磨从发展种子分离。在早期的DPA,富含纤维的RNA可以通过打冷冻胚珠/纤维与玻璃珠分离(Taliercio and Boykin, 2007).从最初的大规模的努力,十多年前开始的(高级研究项目处终端设备等,2004),许多研究表征纤维转录发生。更早些时候,棉纤维转录导致第一次鉴定(假定的)植物纤维素合成酶基因(Pear et al., 1996),这一发现在拟南芥基因证实(Arioli et al., 1998)。利用生物化学、分子和微观技术分析棉纤维表明蔗糖合成酶能够提供底物,如UDP-葡萄糖、纤维素合成(Amor et al., 1995)。低温电子显微镜和生化实验表明,蔗糖合成酶是目前内外棉纤维膜(Salnikov et al., 2003; Brill et al., 2011),这可能与胼胝质(beta;- 1,葡聚糖)和/或纤维素(beta;- 1,4-glucan)的合成。棉纤维的药物治疗表明,纤维素纤维的取向,而不是纤维素合成本身,依赖于微管(Seagull, 1993),这是在拟南芥中证实了活细胞成像(Pare-dez et al., 2006)。对棉纤维的生物物理分析显示纤维素IV(Chanzy et al., 1978),帮助定义alpha;纤维素和beta;纤维素(Atalla and Vanderhart, 1984),并证明与初生壁纤维素和次生壁纤维素分子量较高(Timpa and Triplett, 1993).
次生壁纤维素合成、棉纤维采用植物体的厚壁组织细胞的遗传程序(如木质部管状分子和束间纤维)而矩阵多糖和木质素合成(Haigler et al., 2009; Betancur et al., 2010). 由于棉纤维的形态分为毛这个发现是令人惊讶的。然而,在拟南芥中的厚壁组织细胞次生壁合成相关基因的表观基因上调在棉纤维次生壁沉积的发生(Haigler et al., 2005, 2009; Al-Ghazi et al., 2009; Singh et al., 2009a; Hinchliffe et al., 2010).相反,编码次生壁CesA亚型相关不上调拟南芥叶毛如定量–PCR,微阵列基因,CESA启动子分析:格斯报告基因在转基因拟南芥,和非功能性的次生壁纤维素合酶基因株系的厚毛墙观察(Betancur et al., 2010).这一结果与棉纤维次生壁与融合了传统定义的原生和次生壁组成的几乎是纯纤维素对比一致(包括纤维素、果胶、甘露聚糖、木质素)在成熟的毛细胞壁 (Marks et al., 2008).比较发育调节基因表达组证实传统的初生壁的相关特点的拟南芥叶毛是相对于传统棉纤维次生壁相关的特性,在发病的壁增厚(Betancur et al., 2010). 木质部产生的遗传程序的棉纤维采用的是目前在用这段时间以来变化不大,陆地植物谱系的基础 (Haigler et al., 2009; Hinchliffe et al., 2010; Zhong et al., 2010),也体现在对内之间的基因表达的比较棉物种(Al-Ghazi et al., 2009; Rapp et al., 2010). 厚壁组织细胞,没有叶毛,在棉纤维细胞壁加厚的模式植物拟南芥提供最好的类比。这说明了灵活运用“植物细胞壁工具箱”产生不同的细胞壁特别适合他们的目的(Betancur et al., 2010).
3.棉纤维细胞壁的组成
棉纤维细胞壁成分变化都会动态地发展。在年轻的纤维之间的0和2德:(a)有丰富的果胶,木葡聚糖耗尽外护套的发展;(b)表位丢失的特点是(1–6)-beta;-半乳聚糖(可能包含着阿拉伯糖原位在阿拉伯半乳聚糖蛋白);和(c)一个表位的出现为特征(1–4)-beta;-半乳聚糖,一种侧链的(Vaughn and Turley, 1999; Bowling et al., 2011) 还以2DPA,一个棉纤维中间层的合成(CFML)开始。这薄薄的粘性外初生细胞壁层富含岩藻糖基化和非岩藻糖基化的木葡聚糖和聚半乳糖醛酸没有或相对较低的酯化反应。CFML用来连接相邻纤维组织束,成为高度组织化和紧凑在密闭空间 (Singh et al., 2009a).内棉纤维初生壁包含22%结晶纤维素纤维包围的木葡聚糖和果胶(Meinert and Delmer, 1977; Singh et al., 2009a).糖组与众多的单克隆抗体识别的抗原表位分析植物多糖纤维壁提取物(pattathil et al.,2010)表明,木聚糖和乙酰化甘露聚糖也存在于棉纤维(Haigler et al., in preparation).
基因编码的蛋白,细胞壁松弛蛋白表达与棉纤维伸长过程中纤维长度QTL相关 (Harmer et al., 2002; An et al., 2007).原生壁和次生壁沉积之间的过渡,转录调控,细胞壁降解酶分解CFML这样纤维作为个人发布与初生壁相关糖降低了相关现象,细胞壁水解酶调控,降低果胶和木葡聚糖分子质量为次生壁合成开始(Meinert and Delmer, 1977; Tokumoto et al., 2002, 2003; Guo et al., 2007; Singh et al., 2009a).棉纤维中一次与次生壁沉积的明显过渡阶段。在过渡期间,一个独特的“缠绕”的细胞壁层,在木材纤维S1层相似,放下几天 (Seagull, 1993)。纤维强度大大增加,此时虽然壁增厚是轻薄的(Hsieh et al., 1995; Hinchliffe et al., 2011),这可能是由于不同取向的纤维素微纤丝在缠绕层与原墙。绕组层也可以将原电池和二次电池壁融合在一起。其他的发展变化也发生在过渡阶段:微管排列支持细胞壁中纤维素纤维沉积方向的改变;呼吸速率下降暂时;一些代谢糖的浓度变化;纤维素的合成率随增加至35%(w/w)的纤维素含量;CFML降低;和水平胼胝质峰的棉纤维细胞壁内。在次生壁增厚的发生率增加,纤维素合成了微管和纤维素纤维的纤维轴采用相对陡峭的螺旋 (Meinert and Delmer, 197