TCP基因:一个家庭十年快照外文翻译资料
2023-01-10 16:04:01
TCP基因:一个家庭十年快照
Mar Martı acute;n-Trillo 1 and Pilar Cubas 2
1 Facultad de Ciencias del Medio Ambiente, Campus Tecnolo acute;gico de la Fa acute;brica de Armas, Avda. Carlos III, s/n E-45071, Toledo,
Spain
2 Departamento de Gene acute;tica Molecular de Plantas, Centro Nacional de Biotecnologı acute;a/CSIC, Campus Universidad Auto acute;noma de
Madrid, 28049 Madrid, Spain
TCP基因编码植物特有的转录因子,具有一个bHLH模体可使DNA结合和蛋白质之间相互作用。 TCP基因家族在石松类植物里有五名成员,在真双子叶植物中有大于20名成员。基因的复制和多样化使这个家族生成不同的两个分支(I类和II类),两种分支具有略为不同的TCP域。在这篇文章里,我们总结了我们目前掌握的TCP基因家族进化的情况、他们的规律、这些蛋白的生物化学活性和一些家族成员的生物学功能,值得一提的是它在分生组织中可以控制细胞增殖。了解TCP基因的功能,我们就能把它作为一种工具,用以调节植物的生长模式和在农艺方面调节成我们需要的形态。
一个基因家族的诞生
最先对TCP基因的描述是在1999年,即编码具有所谓TCP结构域的蛋白的一小组植物基因。TCP结构域共有59个氨基酸碱性螺旋-环-螺旋(BHLH)基序,可以让DNA结合和蛋白质-蛋白质的相互作用[1,2]。最初用显然无关的基因编码的4个蛋白质来命名这个结构域,名称“TCP”是来自:玉米墨西哥类蜀黍分支(tb1)[3],金鱼草的环形棘皮纲(CYC)[4],和水稻中有增殖作用的细胞因子1和2(PCF1和PCF2)[1]。tb1基因是一个对驯化玉米[3]的强顶端优势起主要决定性作用的基因。CYC参与控制金鱼草[4]花的双边对称性。PCF1和PCF2是绑定到水稻[1]增殖细胞核抗原(PCNA)基因启动子的主要因子。增殖细胞核抗原(PCNA)基因编码的蛋白参与DNA复制和修复,维持染色质结构,染色体分离和细胞周期进程。十年后,TCP基因家族的全家福需要一个广角镜头才能涵盖这样一个不断成长的家庭。
在不同的植物物种都发现了TCP基因的存在,也已经论证了TCP基因在植物生长发育中的新作用。这些
发现强调了这个特殊植物基因家族在控制植物株型的演变和发展上的重要性。对非模式植物的基因组和转录组进行测序会发现更多的功能未知的TCP基因,它们的功能需要在遗传模型系统中可获得信息的帮助下被部分确定。在这里,我们总结了对这个家族目前所掌握的情况,其进化历史,其蛋白质的生化功能和一些成员在控制植物的形态和演化中所担任的角色。
TCP基因家族的进化
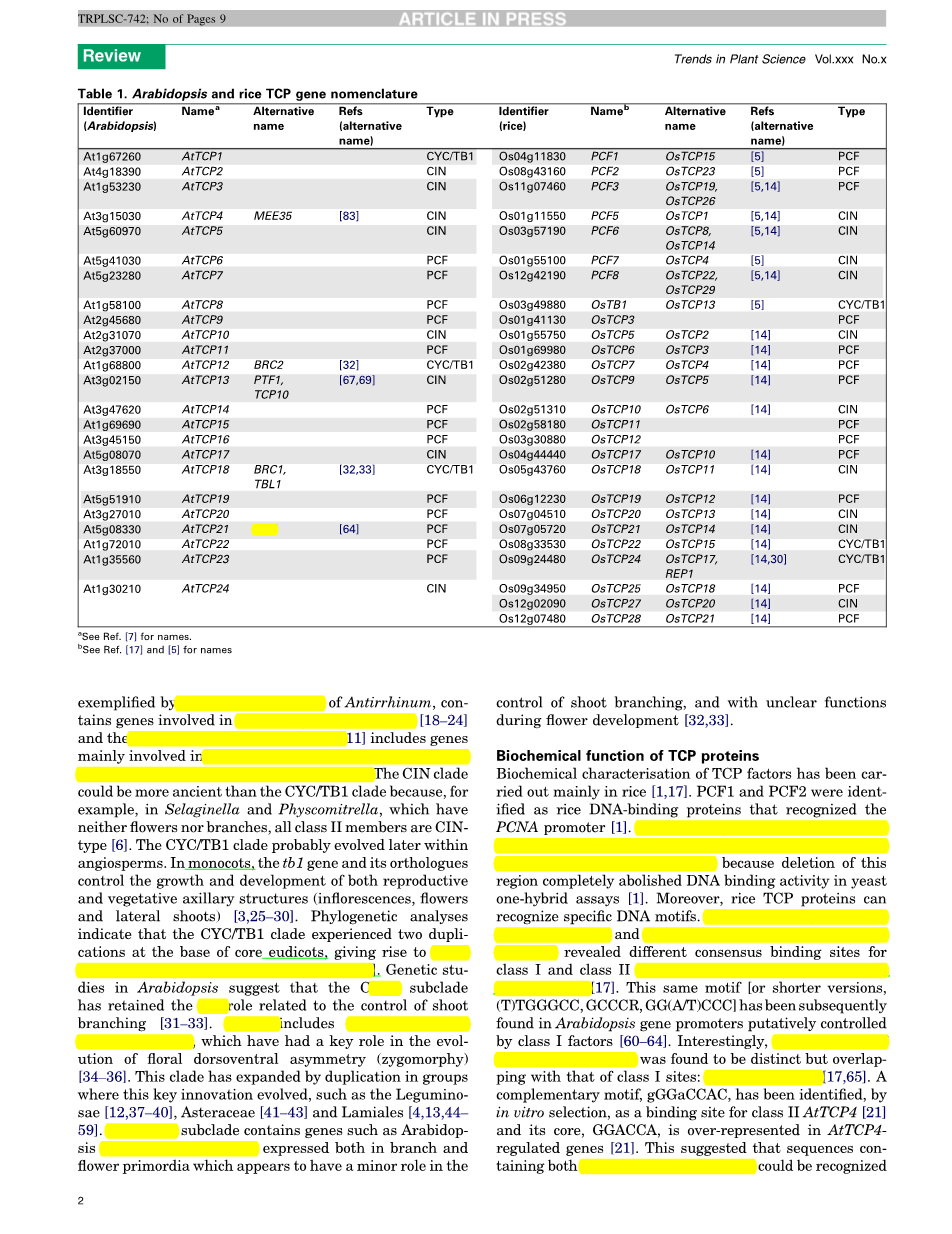
TCP转录因子是古老的蛋白质。虽然他们似乎没有出现在单细胞海藻衣藻中,但是他们在多细胞的绿色藻类中被发现,如鼓藻和轮藻属,在小立碗藓、在蕨类和低等植物苔藓和石松卷柏中也有发现[5,6]。在这些物种中,TCP基因有五到六个成员形成小的家族。通过植物进化、复制和多样化,产生了由数十成员组成的更大的家庭,这些成员存在于裸子植物[5]和被子植物(7 - 16)中。在全基因组搜索时,发现了超过20名成员在拟南芥[7,8]、水稻[5、14、15](见表1完整的列表)、杨树(杨属)[5]和葡萄树(葡萄属)中(材料来自d . Lijavetzky和J.M. Martıacute;nez Zapater的个人沟通)。
基于TCP域的差异可以将TCP蛋白质区分两种类型:I类[17](也称为PCF类[7]或TCP-P类[5])和II类(也称为TCP-C类)[5]。I类包含水稻的PCF1和PCF2,II类包括CYC和TB1(图1和图2,补充材料表S1)。这两个类之间最明显的区别是I相对于II类蛋白在结构域中有的四个氨基酸的缺失。在TCP结构域中的螺旋和环里也发现了两种类型的其他诊断残基(图2)。这个结构域之外的亚类特异的其他的基序,是保守的[2,5,11,14,18]。比如,富含精氨酸的18-20残基的基序(R结构域,图1)[2]和谷氨酸-半胱氨酸-谷氨酸的延伸(ECE基序)[11],这两个基序功能都是未知的,只是知道是II类蛋白的一种。到目前为止,也不能确定哪个类型是原始的TCP蛋白,因为即使绿藻也具有属于这两个不同的组的成员。
在被子植物,I类由一组相对密切相关的蛋白质组成,而II类基于TCP结构域(图1和图2)内的差异也可以进一步细分为两个分支。CIN分支例如金鱼草属植物CINCINNATA(CIN),包含影响侧生器官生长发育的基因[18—24,CYC/TB1分支(ECE分支)[11]包括主要涉及腋生分生组织生长发育,促进花和侧枝生长的基因。CIN分支可能比CYC/TB1分支更古老,因为,例如卷柏属和小立碗藓属,它们既不开花也没有分支,所有的II类成员都是CIN分支类型[6]。CYC/TB1分支可能在被子植物中后来演变出来的。单子叶植物中,tb1基因和其同源基因控制有性和无性腋结构(花序,花和侧枝)的生长和发育[3,25-30]。进化分析表明,在核心真双子叶植物中CYC/TB1的分支经历了两次复制,从而产生三种类型的基因:CYC1,CYC2和CYC3[11]。拟南芥的遗传研究表明,CYC1 子分支保留了与tb1相关的控制分枝发芽的作用[31—33]。CYC2包括CYCLOIDEA-like(CYC—like)基因,这个基因在花的背腹不对称(两侧对称)[34—36]的演变中发挥了关键作用。CYC2分支通过复制来扩大群体,这就是创新进化的关键,如豆科[12,37-40],菊科[41—43]和唇形目[4,13,44-59]。CYC3 子分支包含的基因,例如拟南芥BRANCHED2(BRC2)基因,它在分支和花原基中表达,可以在控制分枝发芽中起到次要的作用,同时在花的发育过程中起到一些不为人知的作用[32,33]。
TCP蛋白质的生化功能
对TCP因子的生化特性的研究主要是在水稻中开展的[1,17]。PCF1和PCF2被确定为水稻中识别PCNA启动子的DNA结合蛋白质[1]。TCP家族的成员都高度保持着TCP结构域中最基本的结构。这个基本结构对于DNA的结合来说是必要(但不充分)的,因为依据酵母单杂交检测结果显示,这个结构的缺失完全废除了结合DNA的活性[1]。此外,水稻的TCP蛋白可以识别特定的DNA基序。随机的结合位点选择实验和凝胶电泳迁移率分析(EMSAs)显示I类和II类因子的结合位点共有序列不同。I类的共有序列是GGNCCCAC[17]。相同的模体[或较短的版本,(T)TGGGCC,GCCCR,GG(A/T)CCC],随后在拟南芥的基因启动子中被发现,假定这些模体是由I类因子所控制的[60 - 64]。有趣的是,我们发现II类的共有结合序列与I类相比是独特但又有重叠的重复:G(T/ C)GGNCCC[17,65]。一个互补结构域gGGaCCAC已经在体外筛选中被确定作为II 类AtTCP4的结合位点。它的核心部分GGACCA,在AtTCP4型基因中过分表达[21]。这表明该序列含有两个位点,G(T / C)GGNCCCAC,可以被这两种类型的蛋白识别,它们可以通过结合TCP位点协同或竞争调节转录[7,17,60]。最近,II类因子CYC已被证明结合I类位点[66],表明序列的特异性并不是绝对的,它甚至允许与TCP因子竞争的可能。
TCP蛋白形成同源和异源二聚体(目前只有对同源二聚体有过描述)[17]。DNA的结合似乎需要二聚物的形成,因为蛋白质的缺失(TCP结构域以外的)可以防止二聚作用造成DNA结合能力的丧失[17]。此外,异源二聚体结合DNA比同源二聚体更有效[17]。因此,可能是将不同的异源组合,绑定到有稍微差异的顺式调控元件,可以识别具有不同的亲和力的靶基因,或调节彼此的活性[7]。这会使由这些因子控制的调控网络在一定程度上变得更复杂。
作为DNA结合蛋白,我们希望TCP作用于细胞核。的确,已经通过免疫沉淀反应的核提取物或绿色荧光蛋白融合物对TCP家族几个成员进行核定位,确定TCP家族的很多成员拥有双或单体核定位序列(NLS)[1,17,32,64,67,68]。相比之下,经描述,融合PTF1/TCP13的GFP被定位到核[67]或叶绿体[69]。这些看似矛盾的结果可能表明,PTF1/TCP13可以针对两个区室。其他的几个TCP因子(AtTCP11,AtTCP15,AtTCP17,AtTCP22和AtTCP23)曾被预言含有叶绿体的靶向肽(cTP)[5,70],增强了这些因素可能控制的叶绿体基因的转录的可能性[69]。
TCP蛋白质与几个基因的启动子上的功能顺式作用元件结合[1,17,21,60-65,69,71]。然而,对于TCP蛋白质控制转录的分子机制仍知之甚少。例如,TCP因子可以激活自己的转录吗?经报道,一些TCP因子在酵母双杂交实验中作为诱饵时可以自激活(M.Martın,F.士和P.古巴,未发表)[17,24],但是在细胞培养共转染实验中,到目前为止还没有测试到I类水稻蛋白质可以被I类位点驱使而反式激活报道基因[17]。因此,至少有可能的是,一些TCP因子本身没有转录激活性,但可以通过与其他蛋白质的相互作用来控制转录。
TCP结合位点常常与其他顺式作用元件相关,表明TCP因子可以充当多聚调控模块的一部分[63,72]。例如,在拟南芥CYCLINB1;1 基因 (AtCYCB1;1)的启动子中,TCP的结合位点与M-特定激活剂(MSA)元件(可控制G2/Mphase-specific的定时表达)的协作[73]可以促进高水平的转录[60]。已经发现,TCP位点与顺式作用元件telo box中AtPCNA2,AtCYTOCHROME C-1,2的启动子和大量的核糖体蛋白基因相关联[61,72,74,75]。 他们在人造启动子方面和telo box共同作用,推动报告基因的表达[61]。可以推断,TCP结合蛋白质参与这些复合物的转录。例如,一个telo box结合因子AtPURa,它在酵母双杂交实验中与AtTCP20相互作用,这表明这两种蛋白质共同作用于核糖体蛋白基因的启动子[61]。另一个TCP因子CCA1 HIKING EXPEDITION (CHE),它结合到的CCA1启动子上,导致基因功能亏损[64],它与TIMING OF CAB EXPRESSION1(TOC1)相互作用,形成一个CCA1的转录激活因子[76]。已经提出,CHE与TOC1隔绝,防止其在CCA1的启动子中与其他转录因子的相互作用[64]。 AtTCP24与Armadillo BTB Arabidopsis Protein(ABAP1)产生不利的相互作用,以调节AtCDT1a和AtCDT1b的转录[65]。最后,金鱼草属中TCP与CUP(TIC)蛋白相互作用,再与NAC域的转录因子CUPULIFORMIS(CUP)结合,可以调节器官的生长界限[77],但其在转录过程中所起的作用尚未得知。
TCP蛋白质作为转录激活因子或抑制因子,但是这似乎并不是由TCP结构域类型确定的:每种类型都包括转录激活因子(i.e. class-I AtTCP20 enhances transcription of CYCB1;1 [60], class-II PTF1/AtTCP13 促进 PSBD的转录 [69]) 或抑制因子(i.e. class-I CHE [64] 和 class-II AtTCP24 [65], 如上所述)。甚至有可能某些TCP蛋白既可以作为转录激活因子也可以作为抑制因子,主要取决于与其他蛋白的相互作用[63]。此外,某些TCP蛋白质的相互作用可能与转录调控不相关。PTF1/AtTCP13促进PSBD在叶绿体中转录[69],已经确定,在酵母双杂交测定中他独立德与含组氨酸phosphotransmitters(AHPS)相互作用,它调解His--天冬氨酸磷酸-elay的信号转导,如调解细胞分裂素的信号转导通路[67]。这就提出了一个可能性,这个因子也可以调控细胞分裂素的信号转导。
蛋白质之间相互作用可以部分被亲水脂性的TCP 结构域螺旋所调节[2],其中一个包含LxxLL的信号(或与之密切相关的fxxLL,其中L是亮氨酸,X是任何氨基酸,和f是一个疏水氨基酸),已被证明它可以调解动物中转录共活化剂与配体的核受体结合[78]。然而,其他部分的蛋白质可以在某些情况下,提高其特异性:ABAP1与AtTCP24相互作用,但并不与和它密切相关AtTCP3 AtTCP5,PTF1/TCP13或AtTCP17因子相互作用,这些因子几乎包含相同的TCP域(图2)[65]。这表明,高度发散的、快速展开的TCP结构域以外的序列,在某些情况下,其功能上的特异性至关重要。相比之下,金鱼草 CUP可以同时绑定TIC 和CYC,这两个不同的TCP蛋白质属于不同的类(图1)[77]。所有这些相互作用,迄今为止已经证明在体外进行,在植物发展体内是否进行还需要证实。
TCP基因在生长发育过程中所起的作用
长期以来一直认为I类基因促进植物的生长和增殖,主要基于水稻PCF1/PCF2和AtTCP20在分生组织表达,而且这些基因是PCNA and CYCB1;1的转录激活子,分别为[1,60]。然而,可以直接证明它功能的证据目前还没有。经分析大多数的I类单突变体没有或有轻微表型缺陷,可能是遗传冗余的结果[60,63,79,80]。而在拟南芥中,AtTCP20融合到EAR抑制域的异常表达(这很可能导致几个I类基因失活),会给表型带来严重影响,这表明I类基因在调节细胞分裂、扩大和分化中有着复
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 9 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[286441],资料为PDF文档或Word文档,PDF文档可免费转换为Word



